ブックタイトル日本結晶学会誌Vol61No3
- ページ
- 35/60
このページは 日本結晶学会誌Vol61No3 の電子ブックに掲載されている35ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
このページは 日本結晶学会誌Vol61No3 の電子ブックに掲載されている35ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
日本結晶学会誌Vol61No3
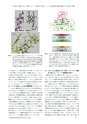
生命科学の飛躍のために一層深まるタンパク質結晶学の役割~シトクロム酸化酵素の精密構造解析から見た現状と課題Cyt.c382383380Lys13Lys87Ser117Asp119 Tyr121Lys8Gln12Tyr105Asp139Helix XHeme a3CcOHeme aCyt.cLeu3811.65A?3.09A?0.33A?Heme a3CcOCyt.cHelix X図14ヘムa3の微小変化によってヘリックスXの構造が変化する仕組み.(Small structural changes ofheme a 3 affects the conformational transition of helixX, which regulates water channel gating.)完全還元型(R)とCO結合還元型(Aモデル)の構造を重ねている.ヘリックスXの中の残基番号はNH-O水素結合が壊れる部分を示している.CcP図15 Cyt,c-CcO相互作用.(Interaction between Cyt.c andCcO.)上図はCyt.cとCcOをCaを結んだ線で表示し,相互作用にかかわるアミノ酸については側鎖の構造も示している.中図と下図はそれぞれCyt.c-CcO,Cyt.c-シトクロムペルオキシダーゼ複合体の模式図である.小球は水分子を表している.Cyt.c-CcOではCyt.c,CcOともに表面の水を保持したままで複合体を形成する.との反発によってP側に動かされる際に,もし水チャネルの弁が開いておればN側への移動も起こる.しかし,A状態で弁は完全に閉じているのでそれが抑えられる.CuBに結合した段階で弁を閉じるスイッチが入り,A状態になった時点で確実に閉じて,プロトンの逆流を抑えている.プロトンの逆流を防ぐ安全性を担保するためにスイッチを入れるタイミングが設定されている.4.8サブAの動きによる弁の開閉制御-ピコスイッチヘムa3に過酸化物やO2やCOなどの化合物が結合するとわずかにヘムが移動する.それが引き金になってヘリックスXの大きな構造変化を起こす.図14ではR状態の還元型とA状態類似構造のCO結合還元型の構造を重ねている.還元型でヘムa 3のビニル基とLeu381のCβの距離は3.37 Aである.COが結合するとヘムa3が移動して還元型構造のLeu381Cβとの距離は3.09 Aに近接するためにLeu381の側鎖がCα-Cβを軸に180°回転すると共にLeu381のCαは1.65 A動く.さらにヘリックスの構造は380-bulgeから382,383-bulgeに大きなコンフォメーション変化を起こして,水チャネルの弁を閉じる.サブAの微細な構造変化によって機能が制御されているのである.まさにピコメートルの精度で作動するスイッチである.日本結晶学会誌第61巻第3号(2019)4.9 CcO:Cyt.c複合体における新しいタイプの“特異62)的で柔らかい”タンパク質間相互作用構造を決めてみて常識では考えられないことに出くわすことがしばしばある.CcO:Cyt.c複合体もその一例である.タンパク質間で複合体を形成する場合,お互いが接触する分子表面の水が排除されて疎水性相互作用によって結合するのが一般的である.ところがCcO:Cyt.c複合体では2つのタンパク質間で最近接Cα間距離は8.2 Aで,これまで報告されている複合体で最も離れたものよりも1.0 A遠く離れている.遠く離れているからといって相互認識が曖昧になっているのではない,お互いに分子表面に突き出た特定の親水性基間で水素結合やイオン対による特異的な相互認識を行っている(図15).相互作用にかかわっているCyt.cの側鎖とCcOの側鎖の温度因子はCcOの主鎖原子と変わらない低さである.一方,相互作用の接触表面積は異常に小さく,タンパク質間に大量の水を含んだ構造になっている.両タンパク質が構造を変化させることなくかつ表面水を保持して会合している.迅速な解離会合を行うに都合の良い“特異的で柔らかい相互作用”である.これまでにこのような報告事例がないが,細胞内では他にも多くあるに違いな179