ブックタイトル日本結晶学会誌Vol61No3
- ページ
- 30/60
このページは 日本結晶学会誌Vol61No3 の電子ブックに掲載されている30ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
このページは 日本結晶学会誌Vol61No3 の電子ブックに掲載されている30ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
日本結晶学会誌Vol61No3
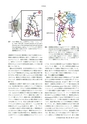
月原冨武P-側CuAシトクロームc弁Mg水クラスターP側CuA水素結合ネットワークCuAe -N側CuB水チャネル弁ヘムaヘムa3H +ヘムaヘムa 3図7CcOの構造(左)と活性中心(CuA,ヘムa,ヘムa3,CuB)の拡大図.(Overall structure of CcO and aclose-up view of the hydrogen-bond network betweenCuA and hemes a and a 3.)シトクロムcから始まる矢印は電子の流れを示す.左図の矢印はプロトン能動輸送を示している.試験管内での水合成(酸素還元)反応では制御されない複雑な連鎖反応が爆発的に進行して発熱を伴う.CcOはどのようにして,発熱を抑制して酸素還元反応を触媒し,効率よくエネルギーを蓄積するのであろうか.構造研究を中心にこれまでの研究成果を総括しつつ今後の課題について述べる.図6は生化学的研究,分光学的研究などによって明らかにされた反応サイクルを酸素還元中心の構造の変化で示している.42)酸素還元中心のFea 3, CuBが還元状態で配位子がない状態のRから順にA,P,F,O,Eの状態が確認されている.酸素分子が特定の決まった順番で構造変化することが,試験管内の酸素還元反応と異なる最大の特徴である.酸素還元を1段階ずつ順序どおりに正確に進めるためには,反応物が過不足なく必要なタイミングで供給されなければならない.そのような高度な制御機能を可能にするためにどのような構造が用意されているのであろうか.CcOは小さな酸素分子の還元反応を触媒するに過ぎないにもかかわらず,プロトン濃度の低いN側(マトリックス側)からプロトン濃度の高いP側(膜間腔側)にプロトンを輸送するために約40 Aの膜を貫通する大きさが必要になっている.反応に必要な電子はP側でシトクロムcから活性中心CuAに渡されて,ヘムaを経て酸素還元中心のヘムa 3に伝達される(図7).一方,プロトンはN側から後述する特定の経路を通って酸素還元中心に送られる(文献42)のFig.9).導線があるかのごとく決まった経路で反応物が供給される.酸素還元に使われる化学プロトンとは別に能動輸送されるプロトンにつ図8N-側能動輸送経路HとMg水クラスターの配置.(Thestructure involved in proton pumping.)経路HはN側の水チャネルとP側の水素結合ネットワークで構成されている.水チャネル,水素結合ネットワークともに弁の役割を果たす構造がある.図中でヘムa3の上部にMg水クラスターがあり,経路Hと水素結合で連結されている.ここにR状態で4プロトン蓄積されると考えている.いては,われわれは哺乳類のCcOでは経路Hで輸送されるとしている(図8).一方,多くの研究者は細菌のCcOの研究に基づいて経路Dで輸送されると主張している.構造に基づいて作業仮説が提供され,さまざまな実証研究によってCcOの働く仕組みの理解が深まっている.4.2ウシ心筋のCcOの結晶化濃縮法で得た最初の結晶の分解能は8A,43)次に塩析法で5 A分解能の回折像を与える結晶を得た.44)さらに界面活性剤decyl maltoside,40 mMリン酸ナトリウム緩衝液で,沈殿剤は当時新たにタンパク質の結晶化に使われ始めたpolyethylene glycol(PEG 4000)を使用することによって,2.6 A分解能の4角板状結晶を得た.45)タンパク質を精製する過程で硫酸アンモニウムによる再結晶化を組み込むことによって,タンパク質試料の純度を高めて2.3 A分解能まで改善した.凍結防止剤45%ethyleneglycolを使用して100 Kで回折実験を行うと1.8 A分解能に向上した.この際に結晶格子の1軸は約3%短くなり,結晶の脱水が起こることが分解能向上の要因になっている.また,凍結防止剤濃度を45%まで高める際に段階的に徐々に変化させることによって,分解能を1.5 Aを超えるまで向上させることができた.46)これは一挙に結晶格子を変化させようとすると,結晶表面と内部では一様に変化しないために結晶にモザイク幅を大きくして,結果として分解能の低下をもたらす.徐々に凍結174日本結晶学会誌第61巻第3号(2019)