ブックタイトル日本結晶学会誌Vol56No3
- ページ
- 51/94
このページは 日本結晶学会誌Vol56No3 の電子ブックに掲載されている51ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
このページは 日本結晶学会誌Vol56No3 の電子ブックに掲載されている51ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
日本結晶学会誌Vol56No3
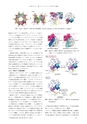
21番目のアミノ酸,セレノシステイン合成の分子機構図7SelAとtRNA Secの複合体の結晶構造.(Crystal structure of SelA and tRNA Sec complex.)tRNA SecのDアームと相互作用し,サブユニットBのコアドメインがtRNA Secのアクセプターステムと,サブユニットCのC末端ドメインがアクセプターアームの末端領域とそれぞれ相互作用していた.今回用いたtRNA Secはセリンを結合していないため,セリンを結合する3’端の位置は不明であったが,アクセプターステムの先端の位置から,本来のセリンを結合した状態では,サブユニットCとDの境界面に存在する触媒ポケットに至ると考えられる.変異体を作製し,活性を調べたところ, N末端を欠く変異体SelA-?NはtRNA Secを結合できなかったのに対し, 2量体変異体(N末端ドメインは欠いていない)はtRNA Secの結合能を保持していたため, N末端ドメインがtRNA Secの結合を担うことが明らかになった. 17)一方,どちらの変異体も試験管内および大腸菌内で活性を失っていたため, 17) SelAの活性にはtRNA Secの結合能だけでなく, 10量体化も必須であることがわかった.4.3.2 tRNA Secの識別機構tRNA Secにはアクセプター-Tアームが通常のtRNAより1塩基対長いという特徴がある. SelAのN末端ドメインがTアームの先端と相互作用しており,アクセプターアームの先端は活性部位に至る(図7).このため,アクセプター-Tアームの長さはSelAの活性に影響を与えることになる.しかし, N末端ドメインがある程度自由に動くことができるため,厳密にアクセプター-Tアームの長さを識別できないと考えられる.実際,アクセプター-Tアームを1塩基対短くすると活性が1/5に低下すると報告されており, 22)これだけでは高い特異性の達成は不可能である.tRNA SecのDアームはほかのtRNAに比べステム領域が長くループ領域が短いという特徴をもつ(図2).複合体の結晶構造ではSelAのN末端ドメインがtRNA SecのDアームと相互作用していた. N末端ドメインの表面とDアームの表面は互いに相補的である(図8).一方, tRNA SecをtRNA Serに置き換えてSelAとtRNA Serのドッキングモデルを作製すると, tRNA SerのDアームの先端付近がSelAと衝突し, Dアームの中央付近はSelAと接触できず,隙間が空いている(図8).このため, tRNA SerはSelAのN末端ドメインとの相互作用が不可能で, N末端ドメインとDアー日本結晶学会誌第56巻第3号(2014)図8 tRNA Secの識別.(tRNA Sec discrimination.)SelAのN末端ドメインはtRNA Serを結合できない.図9 tRNA Secの識別メカニズム.(tRNA Sec discriminationmechanism.)ムの相互作用がtRNA Secの特異性に対し,中心的な役割を果たしていることが明らかになった(図9).Dアームの構造を認識する機構は真核生物/古細菌のPSTKと同じである. PSTKもSelAと同様に,大量に存在するSer-tRNA Serと結合せず, Ser-tRNA Secとのみ相互作用191