ブックタイトル日本結晶学会誌Vol55No6
- ページ
- 26/72
このページは 日本結晶学会誌Vol55No6 の電子ブックに掲載されている26ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
このページは 日本結晶学会誌Vol55No6 の電子ブックに掲載されている26ページの概要です。
秒後に電子ブックの対象ページへ移動します。
「ブックを開く」ボタンをクリックすると今すぐブックを開きます。
日本結晶学会誌Vol55No6
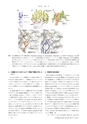
福山恵一,和田啓図4自己触媒反応に伴う構造変化.(Structural changes by autocatalytic maturation.)(a)GGTのwild-typeと自己触媒反応が起きないT391Aミュータントの電気泳動.(b)T391Aミュータントの構造. Wild-typeでは自己触媒反応に伴い大きくコンフォメーション変化をするセグメント(I378-Q390)を黒で示す.(c)T391周辺の前駆体と成熟型のクローズアップ. L-サブユニットのC末端セグメントのコンフォメーション変化に伴い, lidループが所定の位置に来ることができるようになる.(d)前駆体GGTと成熟型GGTにおける活性部位近傍の重ね合わせ. C末端セグメントが移動するに伴い, N411-T416が移動して適切な幅の活性部位が完成する.4.大腸菌GGT以外のGGTで構造や機能は同じなのか?われわれは変わった一次構造をもつGGTに着目した.それはB. subtilis GGTで,一次構造のアラインメントから,多くのGGTでみられるlidループに相当するセグメントがなく, L-サブユニットのC末端セグメントが長い.このような特徴からいろいろな憶測を呼び,構造解析をしてみた. 12)B. subtilis GGTのグルタミン酸結合型では, E. coli GGTのそれとよく似た基質結合構造をとっていた.両者で大きく違う点は前者にはlidループがなく,したがってグルタミン酸は外界から丸見えである.もう1つの違いは, L-サブユニットのC末端はE. coli GGTやH. pylori GGTのようなプロセッシングに伴う大きなコンフォメーションは起こっていなかった. E. coli GGTなどに見られた,酵素反応中に基質が覆われることは必ずしも必要でないのであろう.なお, B. subtilisは高塩濃度で生育できる. E.coli GGTと異なり,このGGTはsurface potentialがNaCl濃度を3 Mに上げても0 Mのときと変化しないことによるからであろう.5.阻害剤の結合様式GGTを阻害する化合物はいくつか知られていた.代表的な阻害剤であるacivicin(図5a)は, Streptomyces sviceusから代謝産物として単離されたものである. 13) AcivicinがGGTのどこに結合して阻害するのか報告されていたが,結果はまちまちであった. 14)われわれはまずE. coli GGT結晶をacivicin溶液にソーキングし,結合サイトを1.65 A分解能で調べた. 18)結果は, acivicinは活性残基であるThr391に共有結合していた.ところが予期しなかったことであるが,電子密度の形状から単純にacivicinのCl原子がThr391のOγ原子に置き換わったようには見えなかった(図5b).われわれはacivicinのC3がsp 3であることから, 5員環内の2重結合が転移したメカニズムを提唱した.その後H. pylori GGTやB. subtilis GGTでもacivicin結合サイト・様式がそれぞれ1.7 Aと1.8 A分解能で調べられた. 16),17)結合サイトはすべて活性残基Thrであったが, acivicinの5員環内の2重結合は転移していないようである. Acivicinの結合様式はGGTによって多様であるようだ(図5b~d).一方,アザセリンもE. coli GGTに結合させ, 1.65 A分342日本結晶学会誌第55巻第6号(2013)